神奇的听觉,仓鸮是如何空间定位的呢?
提到仓鸮(xiāo),我们可能并不熟悉。但如果用另一种称呼——猫头鹰,我们一定立刻联想到这样的形象:

回来吧猫头夜鹰!
应该是下面这样的。。。

一只正经的猫头鹰
其实呢,猫头鹰是一种对鸮形目鸟类的统称,它是鸟纲下的一目,夜行类的猛禽,种类超过130余种。本目鸟类的头部与猫极其相似,故俗称猫头鹰。别名神猫鹰。而我们今天的主角仓鸮是其中的一类,由于面部类似候,又俗称作猴面鹰、谷仓猫头鹰。让我们见识下我们的主角。

仓鸮的凝视
好了,回归我们今天的主题。夜行性动物的捕食除了依靠敏锐的、夜间适应的视觉机制外,灵敏的听觉及空间定位能力也扮演着很重要的角色,尤其在猎物躲藏在遮挡物之后的捕猎行动中。所以仓鸮也因其出众的听觉定位能力很早就受到研究者的关注。
早在1979年,Knudsen和Konishi两位学者就通过行为和电生理记录探究仓鸮的听觉机制。他们发现仓鸮可以通过自身独特的解剖特性,来建立听觉空间的系统的内部表征。他们后续进行了大量的基于仓鸮模型的感觉系统研究。
对于“大脑如何建立起听觉空间的内部表征”这一问题,一直是听觉领域的关注重点。因为迄今为止,我们认识到的内耳声音感受器并不能直接进行空间编码(它只进行声音频率的编码)。在20世纪早期,我们发现大脑是利用间接信息(如:双耳声强差和双耳时间差)来合成听觉空间信息。然而,这一合成计算的过程是如何在大脑中进行的仍尚不明确。
仓鸮经过长期夜间捕猎后演化出特殊行为机制和适应性解剖结构。在自然环境下,仓鸮能够自发准确地将头转向新的目标声源处。而且它们具有非球形的眼睛,意味着不会发生眼跳,因此,通过观察它们的头部的朝向就可以直接判断他们的注视处。Knudsen和Konishi利用这些特征来测量自由状态下仓鸮的定位精度。此外,仓鸮属于一种“不对称型猫头鹰”,因为它们的左右耳的位置在垂直方向上有轻微差异,这种不对称是由前耳瓣和面部皱褶的特殊形态所导致的。
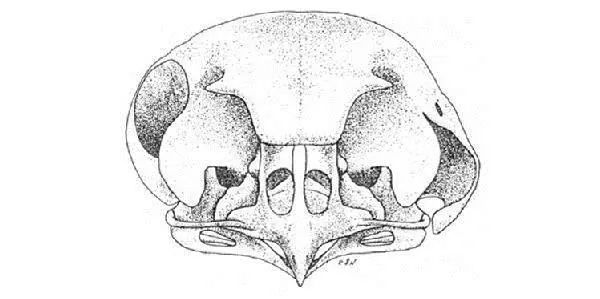
仓鸮颅骨
(从头骨中不难看出左右耳的差异。)
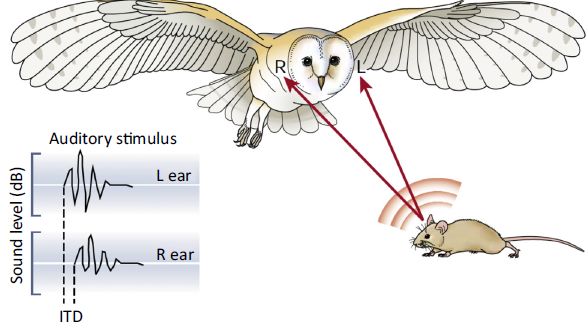
仓鸮即使在完全黑暗的情况下也能根据听觉差异(双耳时间差和双耳声强差)精确地定位猎物
Kundsen 和 Konishi就对仓鸮的水平和垂直方向上的定位进行了评估。结果发现,在水平方向上的声音定位中,双耳时间差(interaural time differences, ITD)和双耳相位差(interaural phase differences, IPD)起到重要作用。神奇的还在于,这种能力几乎适用于仓鸮能够听到的整个频率范围(0~9kHz)。
为什么这些信息可以帮助仓鸮作出方向定位呢?就双耳时间差而言,声音到达双耳的微妙级别的时间差是空间定位的主要依据。对于哺乳动物来说,上橄榄内侧核(medial superior olive,MSO)负责加工双耳时间差。双耳中耳蜗核(ventral cochlear nucleus, VCN)内细胞分别将声音刺激下的兴奋性信号传入到MSO神经元(见图6)。
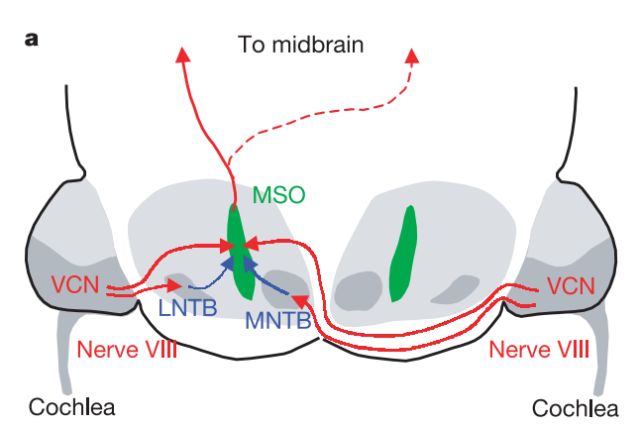
声音信号输入路径
假设在身体左侧有一段声音刺激发生,那么声波会先从抵达左耳的感受器,然后兴奋沿着神经元轴突传递到上橄榄核,右侧在稍晚一点的时刻发生相同的过程。最终来自左侧的兴奋和来自右侧的兴奋相遇,由于左侧兴奋传递时间较长,经过的轴突也较长,故二者相遇的位置在上橄榄核的偏右位置处(见图7)。据此,MSO可以实现对双耳时间差的编码过程。
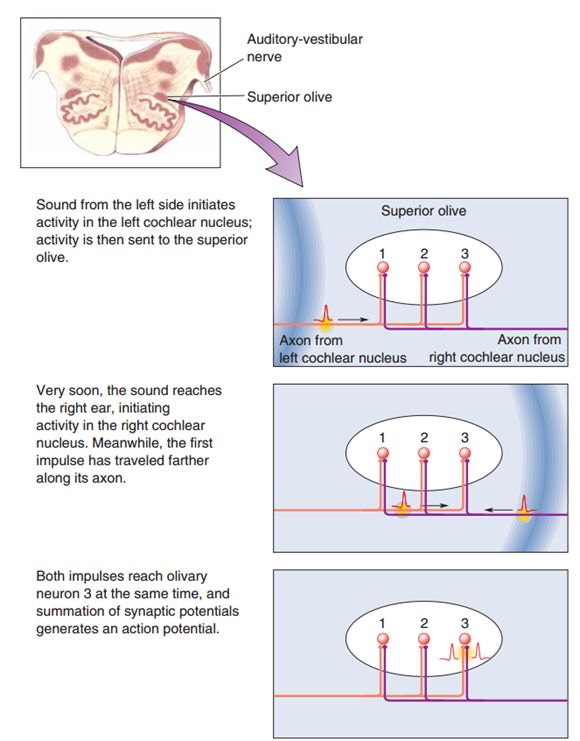
上橄榄内侧核作用机制
相比水平维度的线索,在垂直方向上另一种双耳线索——双耳声强差(interaural level differences, ILD)起到了作用,这被认为与颅骨不对称(双耳水平高度不一致)有关。人类以及一些具有对称颅骨的其他物种能够利用双耳声强差和双耳时间差进行水平方向上的声源定位,尽管双耳声强差不同与双耳时间差,它更适用于更高的频率,其头影效果更强。(头影效果是指颅骨对高频声具有阻拦作用导致的某只耳朵的信噪比上升,从而提高对目标声音的识别)。相反,对于颅骨不对称的仓鸮而言,在水平方向上利用双耳声强差的线索,这也导致了就双耳声强差而言,它的等高线倾向于水平面而非垂直面(见图8右)。相反,对于双耳时间差的等高线则偏向于垂直平面(见图8左)。这些也就成就了仓鸮独特的夜间定位猎物的能力。
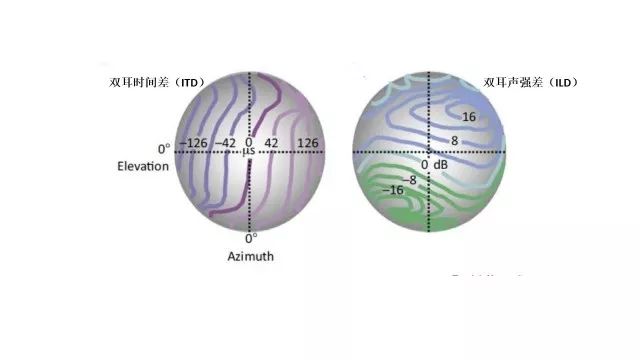
双耳时间差和双耳声强差的等高线图
而后,Knudsen和Konishi又进行一系列针对仓鸮中脑的电生理记录,证明存在独立于声音频率而基于双耳强度差和双耳时间差的声源定位计算机制。Knudsen进一步发现在下丘(听觉中脑)和上丘的外侧核是听觉空间地图和视网膜地图结合的地方。他基于此,还建立了运动控制中发展可塑性和多感官形态整合的模型。
双耳时间差信息是如何在听觉脑干中被加工的呢?大约50年前,Lloyd Jeffress同步检测神经元能基于不同双耳时间差的双边延迟系统补偿的差异来建立方位空间地图。Carr和Konish发现仓鸮的层状核(鸟类延髓内司听觉及声源定位的重要感觉中枢)几乎符合双耳时间差处理的反应模式(这一结论在之后Rubel等人对于鸡的研究中也得到了证实)。这一切都在尝试回答鸟类如何进行声音的空间定位这一问题。此外,还强调地形表征(“地图”)是脊椎动物大脑的一个关键特征。事实上,在20世纪70年代的神经行为学研究中,基于重要感觉线索的地形表征已被提出。例如,Heiligenber和他的同事们发现弱电鱼中脑的神经元敏感性会随着电场性能变化发生敏感变化。在1991年,Heiligenberg 总结到“无论我们在哪里发现行为反应,都是由某个刺激变量的持续调节所引导的,我们似乎找到了一个有序的表示该变量的神经元图谱。”
有趣的是,仓鸮声音定位系统几乎涵盖所有哺乳动物声音定位系统的类别,尤其是在双耳时间差的计算和听觉空间的神经元表达方面。在哺乳动物中,双耳时间差没有排布在计算“地图”中,不论是在进行双耳时间差信息加工的脑干中还是在上丘等高级的加工阶段内。这些差异可能源于哺乳动物和鸟类空间听觉的独特进化起源。总的来说,鸟类和哺乳动物都可以在空间中定位声音,但是使用不同的神经元策略来编码声音的位置。拓宽来说,地形图本身并不是系统处理特定刺激参数的先决条件。事实上,对于需要处理高度复杂和动态环境的感觉信息来说,固定的地形表示法现在很少被认为是一种神经元的“解决方案”。
我们的仓鸮在在神经基础的声音定位的许多方面脱颖而出。虽然在很多时候,鸟类对于听觉任务的结果方式与哺乳动物相似,但至少在某些情况下它们会用不同的神经元加工策略。所以说,仓鸮在某些方面可以算是一类特殊的鸟了。站在进化生物学的角度来看,这是多么有趣的一个事件。它强调了不同的大脑如何整合多样的感觉信息,这种多样性不仅从神经行为学的角度来看是迷人的,而且它还有助于我们更好的理解物种差异以及通过比较去诠释进化论中的原理。
作者:CC
校对:Jacaranda
审稿:庚润
编辑:X君
本文经由公众号:脑人言(ID:ibrain-talk)授权转载,
如需二次转载请联系原作者,
欢迎转发到朋友圈。
ID:Guokr42
整天不知道在科普些啥玩意儿的果壳
我觉得你应该关注一下



