李家洋:为14亿人培育新时代的水稻

未来论坛联合中国科学院遗传发育所,特邀2018年未来科学大奖-生命科学奖获得者、中国科学院遗传发育所研究员李家洋,中国科学院遗传发育所所长杨维才、研究员曹晓风、田志喜,上海生命科学研究院植物生理生态研究所所长韩斌,举办学术报告会生命的礼赞系列之Smart Crops in the Future。下面是李家洋老师演讲的精彩内容:
水稻是我国的主粮,能不能生产足够的粮食,为国民、为子孙后代提供丰富的、有营养的食物是中国乃至全世界都面临的问题。其他方面还包括在生产粮食过程中对环境造成的巨大压力、粮食生产对自然资源的消耗以及食品安全。特别是在中国,我们有接近14亿人口,我们的可耕种土地仅占世界8%左右,而我们的人口却占全球总人口的20%。在这样一个巨大的压力之下,我们怎么样才能够确保足够的粮食生产,而且要高效,也要保护环境、保护资源。
从研究的角度看,那就应该要想怎样才能提高产量,特别是单产的提高。怎么样才能去改善粮食的品质、营养价值。对自然灾害的抵抗能力、少投入化肥、少投入抗病虫害的药以及肥料等等。我们在建立实验室之初,就考虑了要围绕这样一个巨大的挑战或者国家的需求,作为我们的研究方向。要具备一定的特色,我们首先要想办法,要提高粮食产量。如何提高呢?
第一次绿色革命是通过株型的改变提高粮食产量,我们认为在未来新的绿色革命或者是绿色革命的再出发当中,株型可能依然是产量提高的一个关键部分。第二,除了产量,就是如何提高营养品质,使我们的粮食或食物不仅仅是提供能量,也提供生活中必需的营养素。第三是从稳产的角度,能够使作物抗自然灾害、抗病虫害、抗倒伏,确保产量的潜力得到充分地发挥,减少损失。通过对这些性状机理的研究,能够培育出适合不同自然条件的高产的、优质的、稳产的新品种。
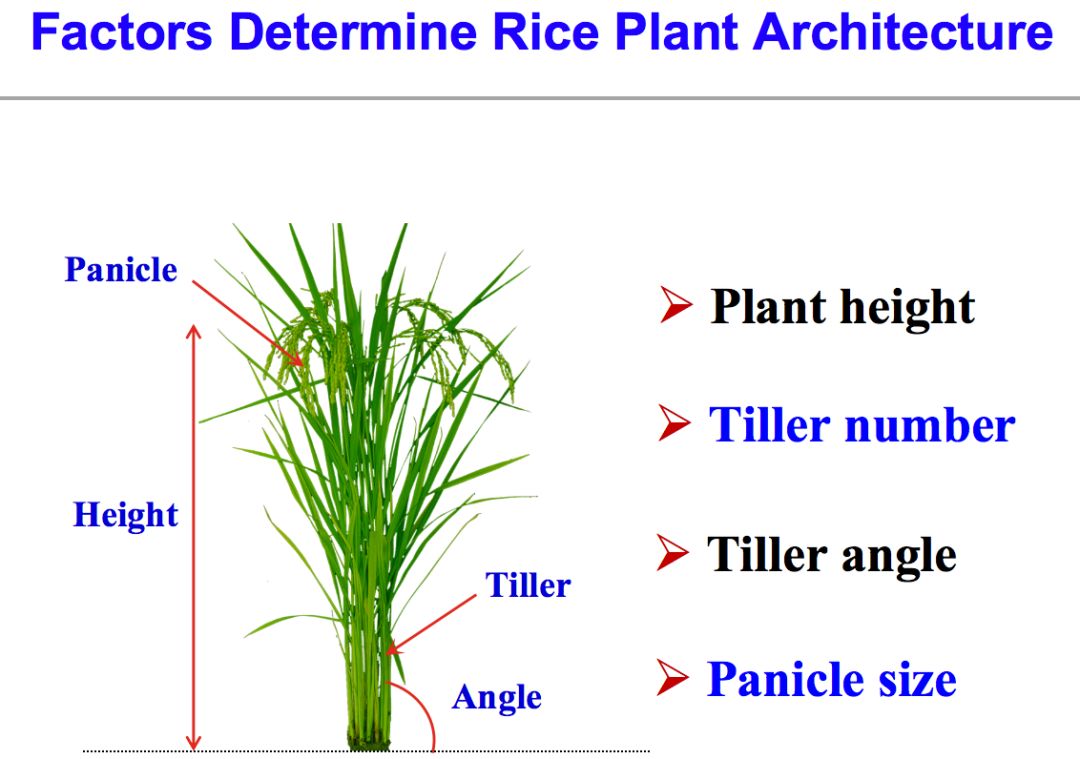
这就是实验室一直为之奋斗的一个大的蓝图。从产量方面,主要是从株型的角度入手。株高是株型的重要构成因素,这是第一次绿色革命的基础,SD1基因的改良带来了半矮杆作物;除株高外,分蘖(分枝)数非常重要,因为分蘖的顶部长着水稻的穗子,分蘖数决定水稻的穗子多少,对产量至关重要。
除此之外,还有分蘖的角度,因为它和光能的利用、植物栽培的密度、对病虫害的影响是直接相关的。再有是穗的大小,与每个穗子的穗粒数密切相关。每亩或者每株作物的穗数,穗粒的大小和穗粒数构成了产量的三要素。从产量角度来说,就围绕这三点进行研究。
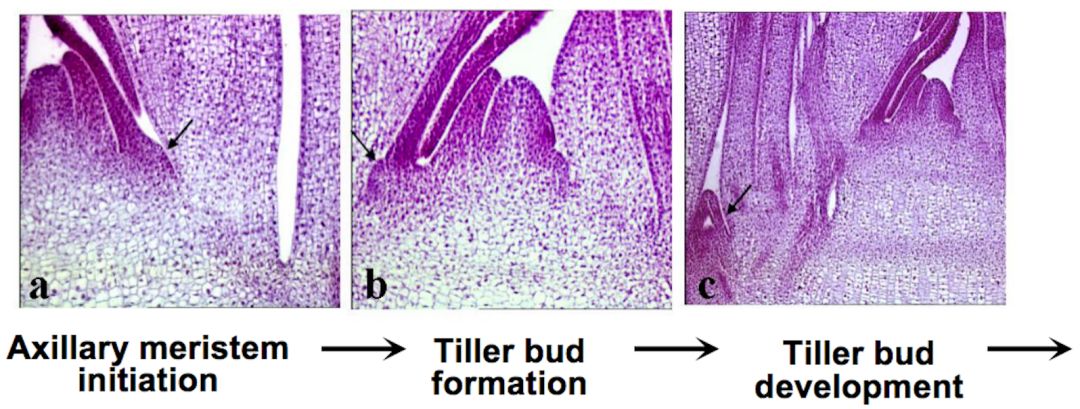
我们先看看分蘖是怎么形成的。有两个主要的步骤,一步是产生分蘖或者分枝的芽,比如在水稻当中,在每一个叶片和茎的夹角之内,产生一个小芽,这个芽长出来形成一个分蘖,分蘖长大之后上面就产生一个穗子。芽的产生是决定性的步骤。第二步就是分裂芽的生长,后面会讲到。

为了理解水稻分蘖芽形成的机理,我们需要材料。我们要找到分蘖数目或者分枝多少改变的材料。我们把它叫做突变体,正常的材料我们叫野生型,Wild-type,用WT来表示。突变体只有一个主穗产生,就是单个秆子,这个单秆材料里尽管叶子依然有,但是在这个地方侧芽完全不能产生。

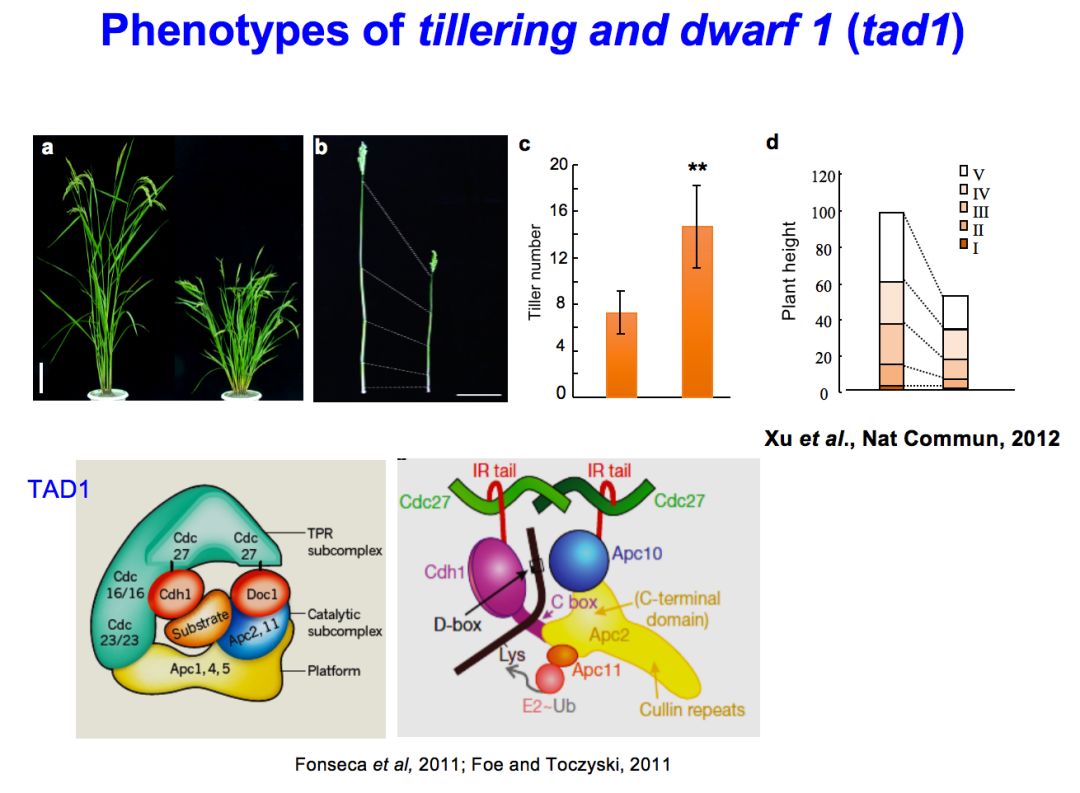
这个基因在2003年的时候发表在《自然》杂志上,是通过基因图位克隆的方法科克隆出来的,这在当时国内是第一次成功的在作物中用该方法克隆基因,也是国际上在主要农作物中克隆基因的新发展。我们一直想知道单秆基因发挥功能的机理是什么,后来又发现另外一个突变体,叫tillering and dwarf 1,简称tad1。
实际上它和moc1突变体反过来,是多分蘖而且矮化的。我们克隆这个基因之后,就发现它编码的是一个重要的降解蛋白质的复合体,叫APC复合体的一个成员,主要负责在这个复合体中识别底物蛋白,进而让底物蛋白降解。这个过程中,它和我们说的MOC1这个基因是互补的,就是说在水稻体内,可以通过物理性的接触结合起来,它们结合起来就可以让MOC1这个蛋白发生泛素化然后降解。MOC1实际上是TAD1的一个底物。
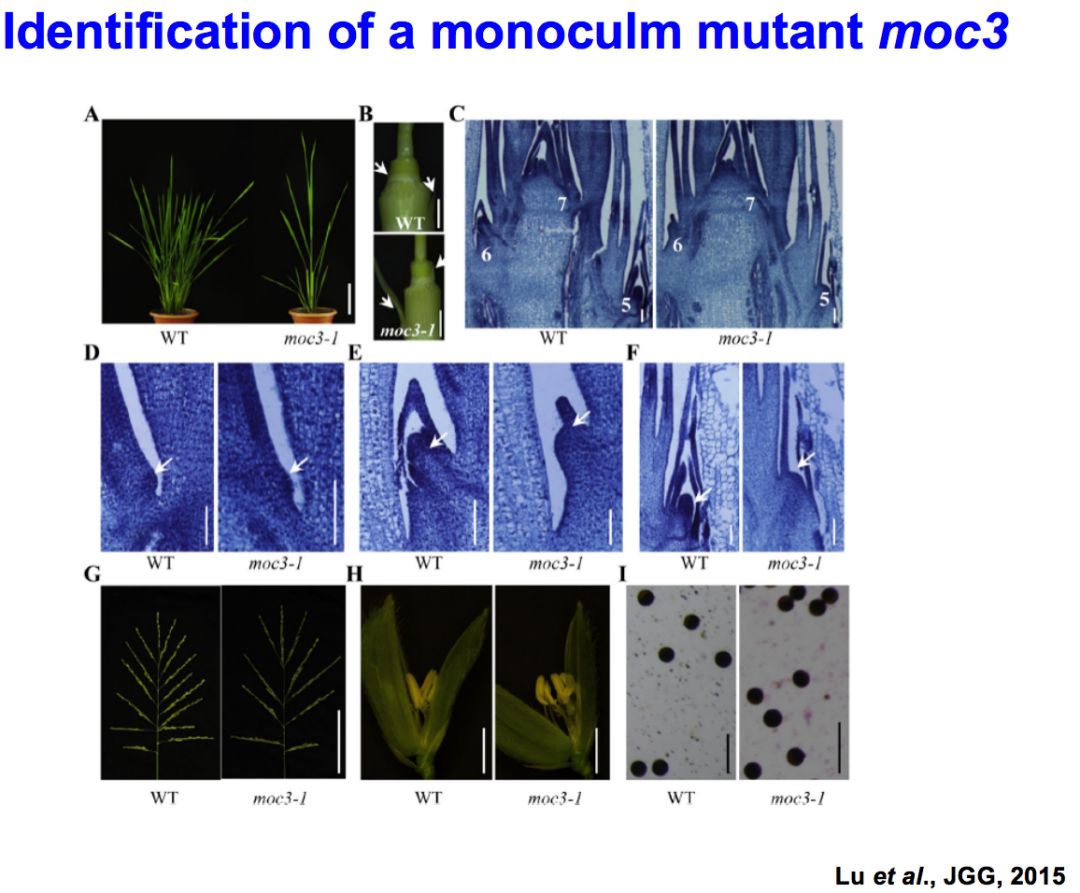
另外一个重要的过程,在分蘖芽形成过程当中,我们还鉴定了第三个比较重要的突变体,叫做Monoculm3 (moc3),这个moc3跟moc1非常相近,也是单秆。而且你可以看得出来,它的顶部的分蘖芽形成的过程也受到一定的影响,但是它的表型比moc1要轻一点,能够形成一定的分蘖芽。
通过一系列的研究,发现在分裂芽形成过程当中,需要一系列基因形成调控网络,实现精准调控,其中MOC1发挥了关键作用。

在实际生产过程中,分裂芽产生后能否发育良好形成一个非常好的穗子可能更为重要,因为很多情况下分裂芽形成后会进入休眠状态。在大约十年前左右,在植物激素或者分枝研究当中发现了一类新的植物激素——strigolactones独脚金内酯。它在棉花、高粱、水稻、豌豆和矮牵牛等植物中广泛存在,但在自然界中含量极其微少,它是由ABCD四个环组成的。
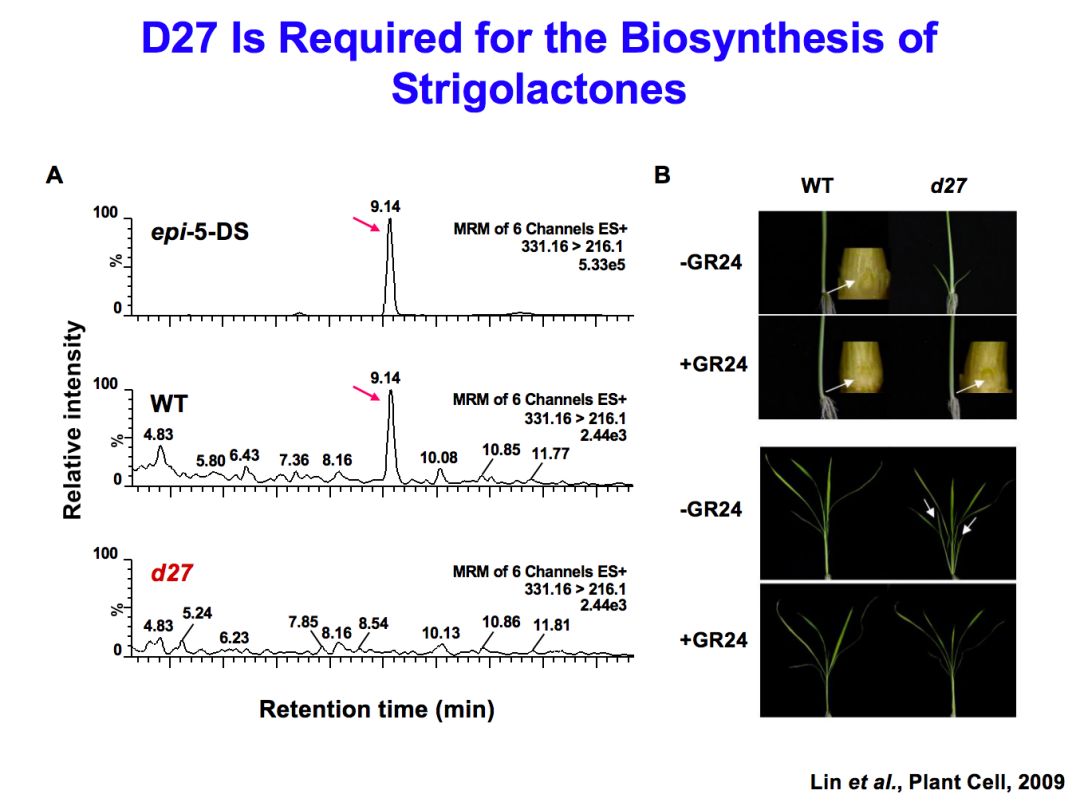
我们首先研究的是它的合成,用的突变体叫的d27。发现与野生的水稻也就是正常的水稻相比,d27缺少一种叫epi-5-DS的strigolactones分子。后来发现D27实际上是strigolactones在水稻中主要的合成途径的第一个酶,非常重要。王永红研究员在这里面起了关键的作用,也做了大量的工作,使这项工作成为研究水稻独脚金内酯合成的一个关键进展。
我们实验室也通过合作,研究它的信号传导机理,D14是它的受体分子,发现它在结构上是形成一个小口袋,这个口袋正好可以把Strigolactones装进去,早期我们发现形成一种共价结合,后来发现ABCD四个环进来后,在D环跟ABC环连接处切开,D环的两端形成共价键,发生构象变化,把信号传递下去。
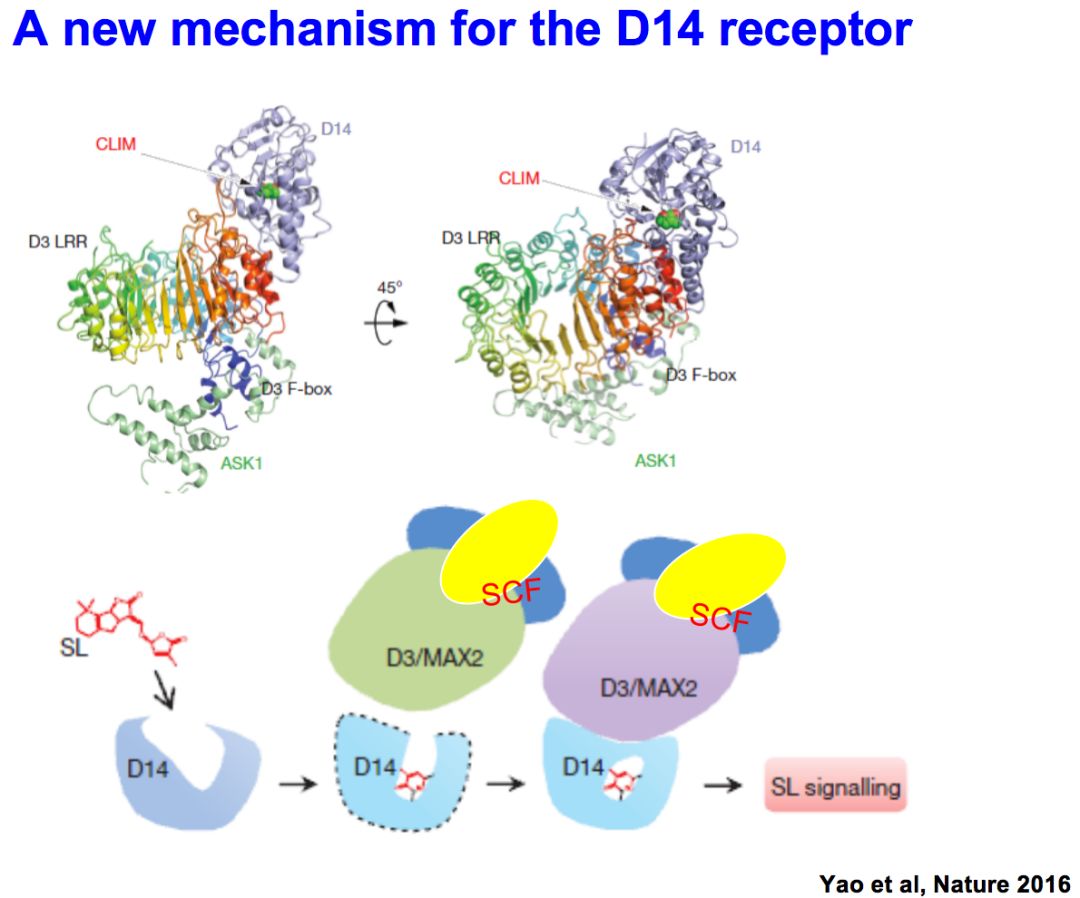
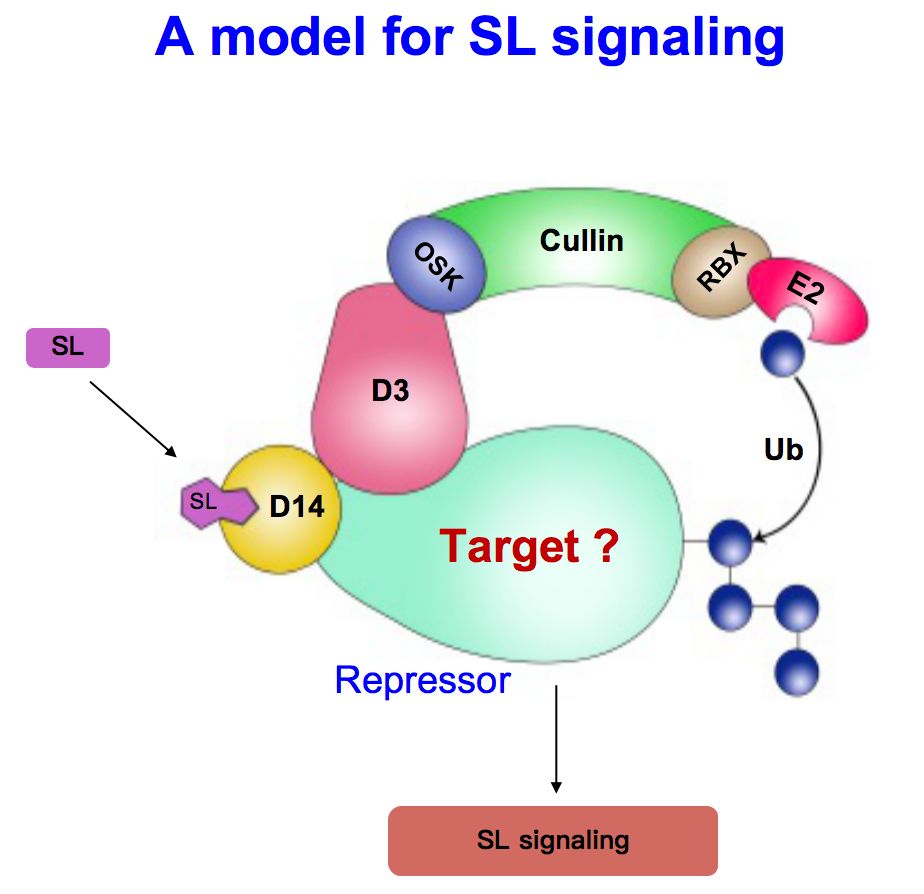
实际上就符合这样一种模式,strigolactones和受体D14结合以后,和SCF复合体结合,使target(底物蛋白)发生泛素化降解,这个target实际上是信号转导的抑制蛋白,抑制蛋白的降解是关键的信号传导步骤。很多在这个领域的实验室都希望找到未知的抑制蛋白。很幸运的是,我们实验室和钱前合作,找到了一个矮生多分蘖突变体,克隆了D53基因,发现突变体中少了5个氨基酸和影响了一个氨基酸的改变,造成了生物学效应。
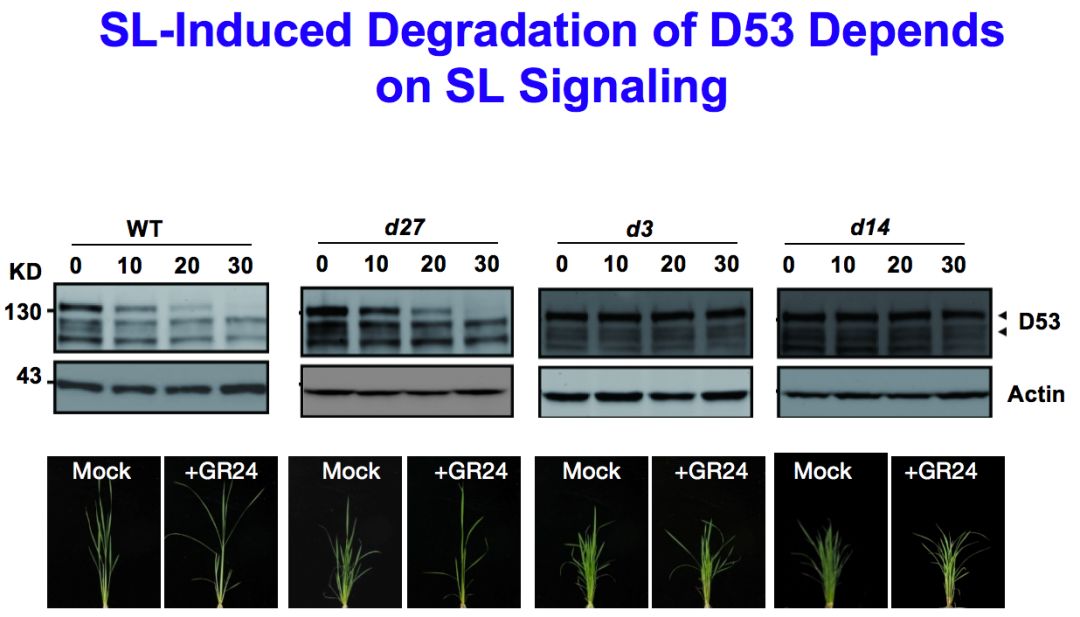
我们进一步做了一系列的检测和分析。比如说D53对GR24是不是有反应,发现在GR24施加之后,D53快速降解,一个小时之内基本被完全降解,d53突变体是功能获得性突变,所以在GR24施加的时候,它的蛋白质不应该被降解,事实上正是这样。通过进一步的分析,发现这个降解是能够被MG132阻断的,说明降解是通过泛素化的形式发生的。
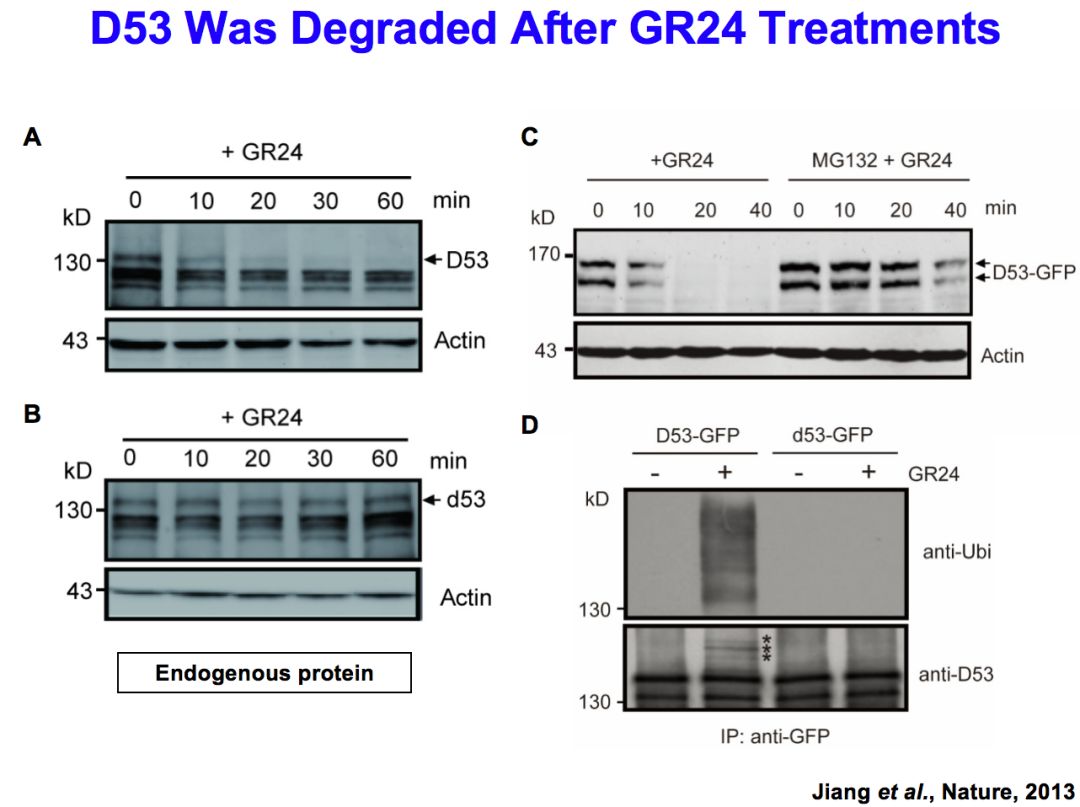
进一步研究证明这是一个信号传导过程,在合成突变体当中,如果使用GR24的话,D53能够完全被降解,但是对于信号传导突变体,比如d3或者它的受体d14当中,D53不能被降解,说明D53的降解需要信号感受和传导的基因。
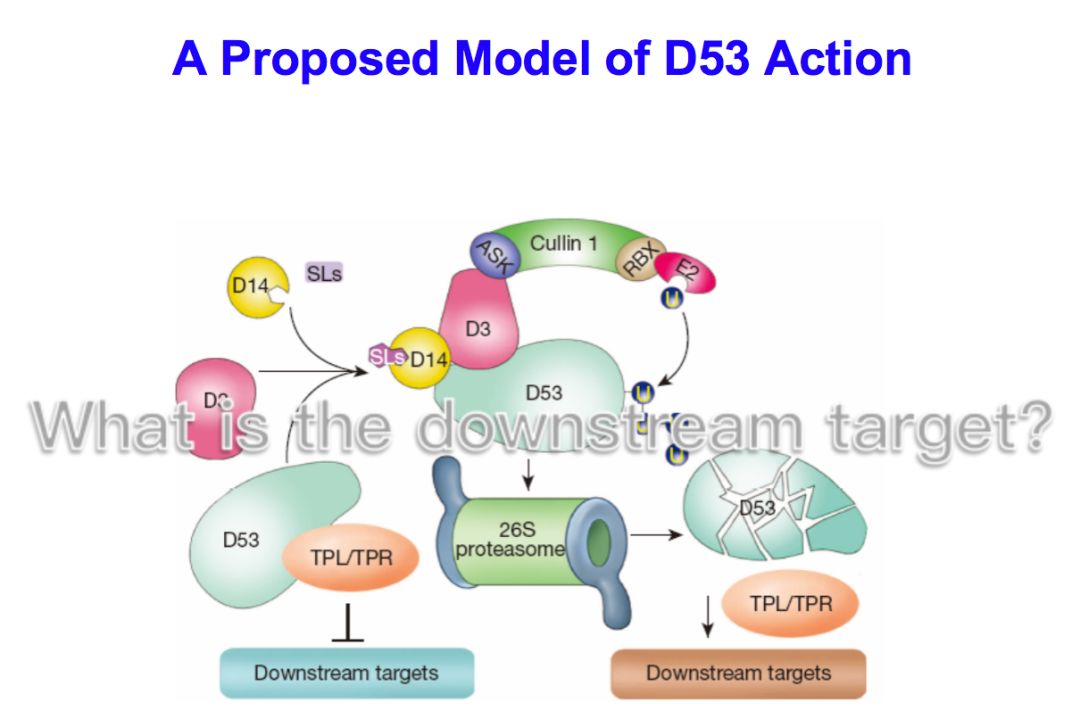
基于以上结果我们提出一个模型,在strigolactones含量比较高的情况下,它的受体D14形成一个巨大的复合体,把D53这个抑制蛋白泛素化,随后通过降解途径把D53降解,下游基因就能够被激活从而发挥作用;在strigolactones缺乏的情况下,抑制蛋白不会降解,它和辅助的抑制蛋白TPL/TPR结合起来,能够非常有效的抑制下游基因表达,让strigolactones信号不能传递下去,从而实现了对分枝的调控。这从机理上解决了strigolactones从合成到信号传导的基本过程。
我们很想知道,独脚金内酯信号下游的基因是什么?实际上我们做了大量的工作。举个例子,水稻专家很早就提出如何能够产生一种最理想的植株形态,得到最高或者最好的产量。这种标准就像有的年轻的人去找对象一样,一定要找到是什么样的一个标准是最理想的、最好的,水稻形态也一样,最好的最理想的一个株型是什么,就是它的分蘖数要适中,不能太少也不能太多,太少了穗子的数目就少。
当然穗子数目通过插秧的密度可以进行弥补,另外穗的大小很重要,要是大穗,当然粒子也要相对大。同时茎杆也要特别强壮,不能倒伏,如果茎秆不强壮的话,有雨、风或者施肥过量都会造成严重的倒伏,另外根系要发达。

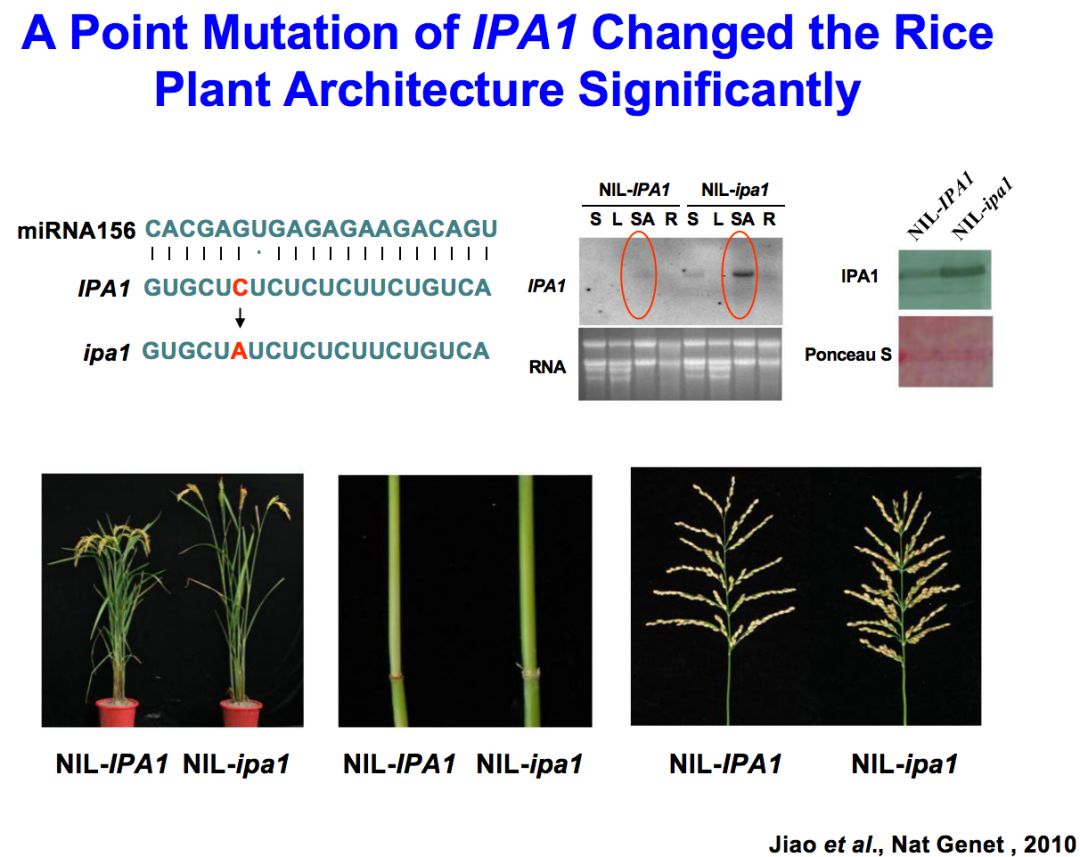
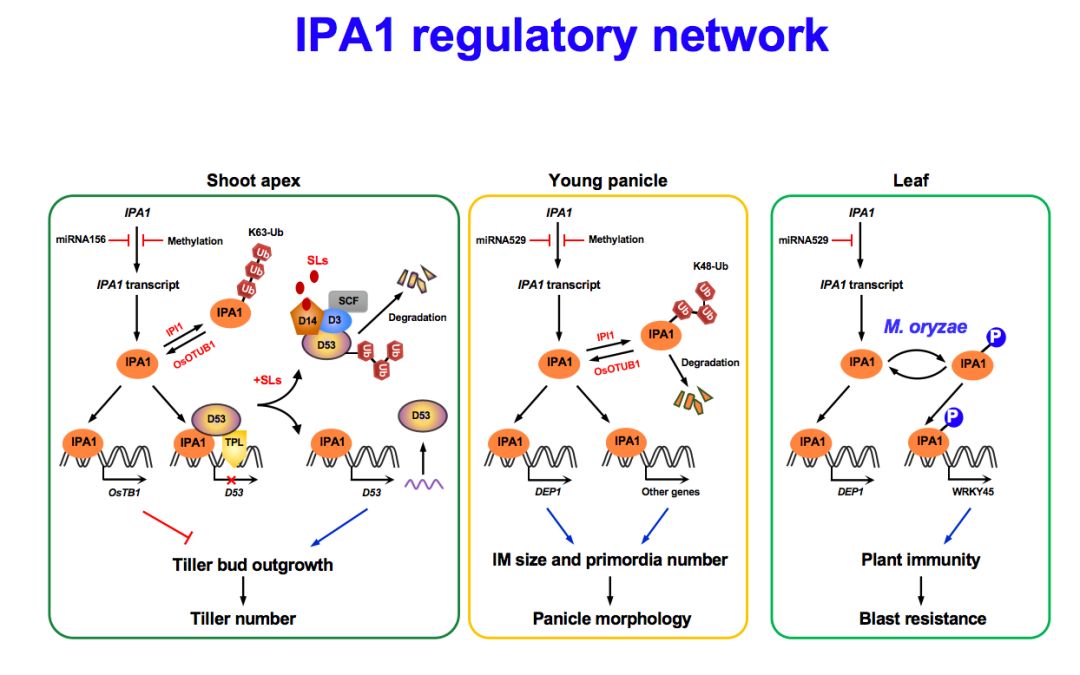
我们发现了一个突变材料,它的很多性状非常符合育种家们期盼的特征,所以我们就把它命名为理想株型1号。把这个基因克隆之后发现它含有两个非常重要的结构域,一个结构域叫SBP,是DNA结合结构域,另一个结构域被miRNA156和529识别进而发生剪切,是调控该基因表达水平的非常关键的机制。
我们这个突变体中理想株型的性状是显性的,和其他材料杂交以后,它们的后代依然有这样的性状,在生产上是比较好用的,这也是我们非常高兴的一个地方。
这个基因实际上发生单个核苷酸从C到A的变化,导致了一个氨基酸的改变。但实际上更重要的是这个位点正好是miRNA156的识别位点,核苷酸的变化使得miRNA156和529对mRNA的识别效率大大降低,剪切效率降低。最终使蛋白质含量增加,因此是获得了功能的突变。
这个基因我们发现它对分蘖数来说是一个负调控因子,如果把这个基因剔除了,失去功能的突变体表现是分蘖增多、株高变矮。如果在miRNA那个位置进行一次改变,使它不被切割而获得功能,突变体的分蘖减少、株高适当增加。
同时,我们也进一步证明了它不论是失去功能或者获得功能的时候,对strigolactones都是不敏感的。进一步研究发现IPA1和D53上实际上是可以互作的。IPA1是作为一个转录因子在起作用,证明了它作为activator发挥功能。这样我们就发现D53能够抑制IPA1的活性,使它对下游基因的转录激活被抑制,改变D53的蛋白水平,当100%是IPA1蛋白的时候,它的下游基因表达很强。
如果D53的量占到20%,下游基因表达就弱了,40%的时候更弱。如果是100%的D53,表达就消失了,所以D53能够抑制IPA1对下游基因表达的激活。
换一个角度看,IPA1表达的积累也可以抑制多分蘖的表型,不同的突变形式都证明了它具有这一作用。IPA1可以结合到Promoter D53上,进而调控D53的表达。可以看到在GR24处理之后,D53的表达是可以提高的,转录的相对水平增加60%左右,还是非常显著的。

最后,我们可以给出这样一个模型,在有strigolactones的情况下,D14结合它,D53蛋白被降解,IPA1受到的抑制解除,使D53这个基因能够表达,其他下游基因也能得到表达。但是如果没有strigolactones的话,D53和IPA1可以紧密结合在一起,就把它们后面的转录抑制起来,调控分蘖的多和少。
可以总结一下,在水稻分蘖形成过程中,可以通过TAD1、MOC1和MOC3这样一个网络,对分蘖芽的形成进行调控。在分裂芽的进一步伸长过程中,strigolactones是一个重要的调控激素,它的合成基因都起到影响作用,在信号传导过程中形成一个比较复杂的网络。我们知道,IPA1是一个非常神奇的基因,它影响了很多的方面,在其中一个方面,对分蘖多少的调控是通过与strigolactones的互作关系来实现的。当然它对茎秆粗壮性、穗大小等可以通过其他途径调控。
这个IPA1基因能够有效的提高单株的产量,实际上也可以通过调控种植密度来影响总的产量。我们进一步和上海的何祖华研究员合作,他们做了大量的工作,证明IPA1有非常强的剂量效应,它的表达量非常重要。如果这个基因的表达处于一倍、两倍、三倍、四倍,实际上分蘖的多少、株高、穗的大小是有一个变化的。
这个变化通过不同的基因型材料,比如我们刚才说的miRNA156位点发生突变的材料,还有一个新的材料,在IPA1的Promoter(启动子)区域,有一个大片段插入,影响甲基化水平,让它的表达量增高,这个增高是比较恰当的,能产生高的产量。

最后就给出这样一个比较新的理论上的预测和判断。IPA1在恰当的表达量时达到最高的产量,一定要关注它的时空上的表达,因为IPA1在不同时空表达产生的后果是不一样的。
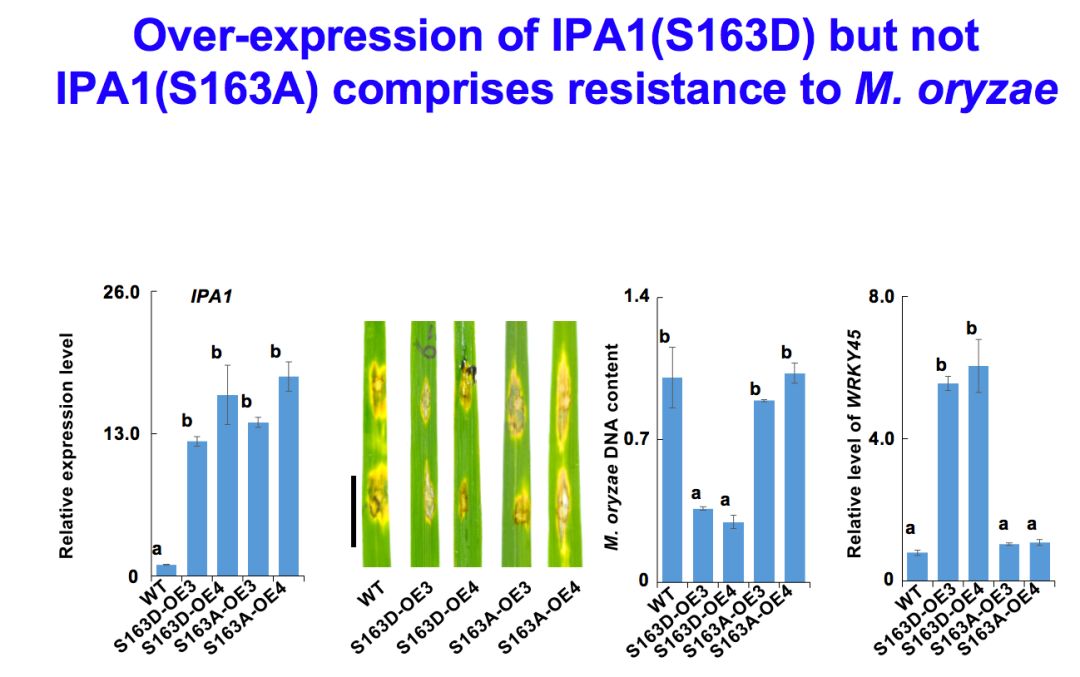
下面我们再稍微说一下IPA1除了产量之外还有另外一个重要的方面,就发现在这样一个突变体的背景当中,它的病斑的大小比较大,过表达IPA1的材料中病斑就非常小。你可以看到,如果是RNAi造成IPA1的表达减少的话,病斑就非常大了,菌株的量也呈现这样的趋势。实际上IPA1蛋白是非常重要的,一旦稻瘟菌感染,它的IPA1蛋白质就被磷酸化了,磷酸化它就能够发挥功能。
我们进一步发现,之所以能够产生这样的效果,就是IPA1有一个位点能够发生磷酸化,只有在S163D这个位置的组成型磷酸化突变有效果。但是在其他位置突变就不行,所以在这个位置的磷酸化起到关键的作用。
我们后面还进一步说明,它是激活了另一条途径,叫WRKY45。实际上IPA1可以提高产量,同时也可以抗稻瘟病。最后用这个图做IPA1的一个总结,在茎秆结构表达的时候是影响分蘖的多少,就是分枝多少,如果在穗上表达,是影响不同的泛素化的变化,它就是影响穗的大小。
但是在叶子里面是高表达的时候就产生抗稻瘟病,所以不同的时空上的表达是极其重要的。稻瘟病没有48小时之后,不侵染它的时候,激活的WRKY45的抗病途径就关闭了,促进高产的途径激活,这样的话就完成了这样一个模式。
下面讲一下关于品质和育种方面。在品质方面,主要讲一下Starch(淀粉)合成的东西,因为我们吃的大米90%都是淀粉,所以淀粉非常重要。这个工作最初的时候我们就在考虑,可能要开创一个新的方法,我们当时刚刚来的一个研究生,田志喜研究员。
他就做了一个比较有挑战性的工作,当时我们不敢做这个,因为水稻好吃不好吃,品质这个事儿,这个实验在国际上做的特别多,特别复杂,怎么把它搞清楚,究竟哪些基因是影响品质基础的,要把它搞清楚。我们就问他能不能做这个挑战工作,他非常勇敢的承担了这个课题,因为这个可能是关系到毕业的问题。他通过查文献,在动物上已经开始做了,比较成功。
在植物上可能有一例做开花的,可以通过一定的关联分析的方法。既然植物上有一例能够做开花,那我们凭着这个现象也是可以的,最后就做了。这样我们和钱前、顾老师,也在韩斌研究员的帮助下,选了70多个不同的研究品种,一部分是籼稻,一部分是粳稻,因为粳稻要相对好吃一点,决定它影响的有四个因素,直链淀粉和支链淀粉的含量,我们把它们归结为直链淀粉的含量(AC),还有胶稠度(GC)和糊化温度(GT),每一个品种都进行测定。
测定完以后,我们就对18个淀粉合成的基因进行了分析,这在十几年前的时候还是非常困难的,要分析编码序列、非编码序列,平均每一个基因大概有10个KB的长度,那时候很难,用PCR扩增出来,选了70个不同的品种,每一个品种有20个基因。最后他还是非常有创造性,说70个不能都做,做14个核心品种,对核心的品种进行基因测序,其他就进行一部分的测序、分析。

最后就给出这样一张图(上图),这张图一看到之后就非常明确了。图中间的基因是表型控制的相关基因,直链淀粉和支链淀粉含量的主效基因,还有胶稠度的主效基因都是Wx基因,糊化温度GT当中,Wx是一个次要基因,这才是一个主效基因,放在核心中间的位置,还有其他的一些基因。这个图出来以后就把淀粉食用性基本品质搞清楚了。我们做了几十个不同的品种,对这些主要基因的独特性都找出来了,找出来以后就拿到育种上去。
总之,有了这么多的重要基因,和其他的克隆一系列的关键基因,我们都可以把它作分子标记,然后去杂交,根据我们在不同的地区,比如在长江中下游,还是在东北,还是在西南,在不同地区的自然条件,对于类似当地消费者的习惯,把重要的有效资源在做之前就研究清楚,然后找受体、供体,做很好的杂交,再到地里去选育。
比如“嘉优中科1号”连续三年,我们在江苏沭阳万亩田的试验面积上,都是农民自己种的,平均产量都在840公斤左右,有的年头可能高一些,比如今年大概到900多公斤,特别好的地方超过1000公斤,连续几年都很好。
同时我们用这样的思路,在东北五常做育种改良,这是当地的品种(稻花香),这是我们的“中科804”,这是绝对抗倒伏的,今年在大风大雨的情况下,当地的品种80%以上都倒伏了,这样的话“中科804”这个品种就特别好,所以丰产性、抗倒是关键。

这是我们所有走过来的同事、师生、科研人员,以及毕业的这些人员。我在报告当中,也对主要的一些人做了介绍,但实际上还有很多其他的人,比如刘贵富研究员、余泓、王冰等都做了大量的工作,过去毕业的学生也做了很多贡献。我们合作者也有很多,韩斌教授、钱前,还有王(永红)老师,过去我们是一个课题组,也是同事,也是重要的合作者,我们也感谢国家科技部、国家基金委、科学院和研究所给予的大力支持,也感谢今天上午所有作报告的同事。谢谢大家!

特别鸣谢:感谢中国科学院遗传发育所的王冰老师对本文的校对。
---END---






