叶片的“变形记”
叶片是最基本的植物器官。大多数高等植物用叶片作为“太阳能电池板”来接收阳光。扁平宽大的形态有利于增大光合作用的面积,自然而然成为了叶片的特征。如果你的眼睛足够好,你可能会发现小叶片刚刚长出来的时候并没有那么扁平宽大,而是逐渐展开的。
那么,叶片是如何展开的呢?
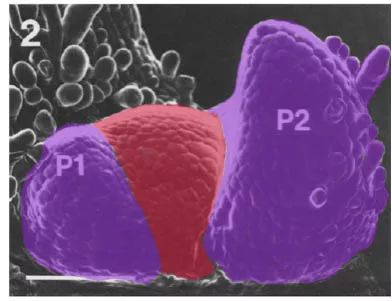
扫描电子显微镜下的烟草叶片原基
(Poethig and Sussex, 1985 Planta)
叶片是在茎尖形成的。茎尖的生长点称为分生组织(上图红色区域),是一团有组织的干细胞。刚刚形成的叶片也是一个小圆包(左侧紫色区域),和生长点差不多。但是,小叶片很快开始变形,分化出背-腹两面。两侧不对称生长,成为包向茎尖的半弧形(右侧紫色区域)。面向茎尖一侧称为近轴面,将来发育为叶片的背面,朝向太阳;另一侧称为远轴面,将来发育为叶片的腹面,朝向地面。
叶片的背面比较光滑,有比较厚的蜡质层,并且颜色更翠绿一些;而相对的,腹面就要粗糙一些,角质层薄一些,进行气体交换的气孔更密集。这些特化非常适应于叶片的光合作用和蒸腾作用:叶片背面负责吸收光能,进行光反应,需要更多叶绿素;叶片的腹面进行大量气体交换,需要更多气孔。

叶片的背-腹两面往往有明显差别
形态学和遗传学的研究发现,叶片展开的第一步就是对称原基的变形,变形后产生了有差异的背-腹两面。背-腹两面之间的边界区也随之产生。边界区向两侧伸展,从而使叶片展开。
简单来说,叶片展开需要两步:变形、展开。那么小小的叶片原基如何知道自己该这样发育呢?中科院遗传发育所最近的工作给出了一些线索。
第一步变形首先有化学信号作为指令。一方面,叶片发生的区域貌似均一,实际表达的基因并非如此。叶片原基形成之时,背-腹轴面就已经隐含其中。另一方面,茎尖的干细胞团帮助叶片原基认清方向。通过对番茄的茎尖进行成像追踪,我们发现经典植物激素生长素从叶片原基向茎尖定向运输,使得叶片的背面生长素浓度低于腹面。基因和激素一起作为化学信号,向叶片原基不同区域的细胞给出了不同的指令:成为背面或是成为腹面。
得到了指令的细胞如何将命令转变为生长呢?答案可能在于细胞壁介导的生物力学过程。中科院遗传发育所最近的一项工作发现,背-腹基因与生长素指令都能够影响叶片原基细胞的细胞壁硬度。植物细胞内部具有很高的压力,细胞壁约束细胞。
通俗地说,每个植物细胞都是一个高压气球,细胞壁就是气球皮。假定内部压力恒定,气球(植物细胞)的大小取决于气球皮(细胞壁)的弹性系数:皮(壁)越硬,气球(细胞)越小。叶片有众多细胞,这些细胞之间不会相对位移,因此叶片的生长需要这些黏连在一起手拉手的细胞相互协调。
根据细胞壁硬度,叶片原基不仅存在背、腹区域,还有一个中间区域。它们的硬度随着叶片生长动态变化。在计算机中,用以上的条件模拟叶片原基细胞,再加入细胞壁的硬度变化,我们就能获得与实际类似的叶片截面变化。因此,细胞壁介导的生物力学变化可能把基因、生长素的命令转化成了三维形态的变化,使叶片原基实现了背-腹不对称的变形。

下一步就是叶片的伸展。刚才提到了背-腹面之间的中间区。这个区域表达一类特殊的 WOX 基因,它们能够促进细胞不断分裂,通过提供新的细胞促进叶片的展开。如果没有中间区的 WOX 基因,叶片只是一条细丝。中间区 WOX 基因的过度表达则使叶片无限生长。
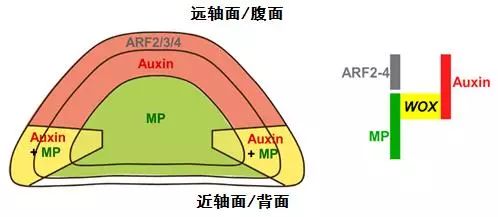
生长素(Auxin)及其下游信号分子(MP)激活 WOX 基因指导叶片展开
那么,这么重要的中间区 WOX 基因又是如何激活的?为何只在中间区激活?
中科院遗传发育所最近的另一项工作发现是生长素与基因的时空特异分布为叶片又划定出了中间区这个新的区域。前面提到了叶片原基腹面富集生长素。生长素是一种简单的小分子,它起作用需要通过受体和下游信号分子。
如果说生长素是一支令箭,那么下游信号分子才是执行命令的士兵,开启了特定的基因表达和蛋白质合成。生长素下游信号分子的分布恰好与生长素的分布接近互补,在叶片原基中主要分布于背面。这样一来,背面有令箭,却无人执行;腹面有士兵,却得不到命令。最终即有令箭也有士兵的只有二者交汇的中间区。
生长素所布置的一个重要命令就是激活 WOX 基因的表达。如此这般,背-腹的不对称性转化为了中间区的界定和展开。看似自然而然的叶片展开,其实是诸多基因、激素协调表演的结果。
来源:中国科学院遗传与发育生物学研究所




