研究发现m6A修饰增强长时记忆形成效率
长时记忆的形成是哺乳动物适应环境变化、智力发展所必需的,对于人类社会活动尤其重要。虽然以往研究已经揭示了一些与记忆形成相关的基因,但关于记忆形成效率的调控因素与机制仍然未知。N6-甲基腺嘌呤(m6A)是哺乳动物细胞中 mRNA 上最为普遍的修饰,自2012年以来逐渐受到广泛关注。已有多项研究表明 m6A 是哺乳动物胚胎发育和多种组织器官形成所必须的,并且参与肿瘤发生等多种重要生物过程调控,但尚不明确 m6A 是否参与了记忆形成调控。
中国科学院遗传与发育生物学研究所王秀杰研究组联合中国科学院北京基因组研究所杨运桂研究组利用胚后敲除 Mettl3(m6A甲基转移酶)的小鼠模型系统研究了 m6A 修饰在成体动物高级神经系统中的功能。利用多种动物行为学检测并结合突变-回补等实验发现:m6A 水平正向调控了小鼠的长时记忆能力,成体海马体神经细胞中缺失或增加 m6A 修饰(敲除或过表达 METTL3)的小鼠分别表现出明显的长时记忆能力下降或上升。除长时记忆能力受损外,这些 m6A 缺失小鼠的生理状态、大脑结构、运动能力、探索行为、趋触性、焦虑水平甚至短时记忆能力均未受到影响。有意思的是,在给予足够训练的前提下,上述由 m6A 丰度决定的学习能力差异消失,所有小鼠达到了同样的学习效果。说明通过 METTL3 调节 m6A 丰度可以影响小鼠学习速度的快慢,但缺失 m6A 虽然导致长时记忆形成变慢,但在给予足够训练的前提下,并不影响最终记忆效果。机制方面,m6A 缺失并不影响神经元的基本电生理特征以及突触短时程可塑性(short-term plasticity),而是减弱了突触长时程增强效应(long-term potentiation)。虽然 m6A 修饰在学习过程中动态变化,但被修饰基因的 mRNA 丰度并不受 m6A 影响。进一步研究发现,m6A 修饰正向调控了神经元兴奋状态下特异表达基因的翻译效率,从而影响了动物的长时记忆形成。
研究进一步发现,野生型小鼠的水迷宫学习能力与其海马体的本底 METTL3 蛋白丰度呈现中度正相关的关系,能在短期内完成水迷宫测试的小鼠的海马体内 METTL3 蛋白丰度要显著高于不能完成测试的小鼠,提示 m6A 修饰丰度的差异可能是造成个体学习能力差异的原因之一。值得关注的是,上述差异在长期训练后的行为学表现分组中并不存在,说明记忆能力的差异可以被多次学习巩固所弥补。由此可见,“勤能补拙”具有生物机制方面的合理性。
该成果于10月8日在线发表于 Cell Research 杂志上(DOI:10.1038/s41422-018-0092-9)。王秀杰组博士生张泽宇、副研究员王猛为该论文的共同第一作者,王秀杰与杨运桂为共同通讯作者。该研究得到科技部、国家自然科学基金委和中科院战略先导专项的资助。
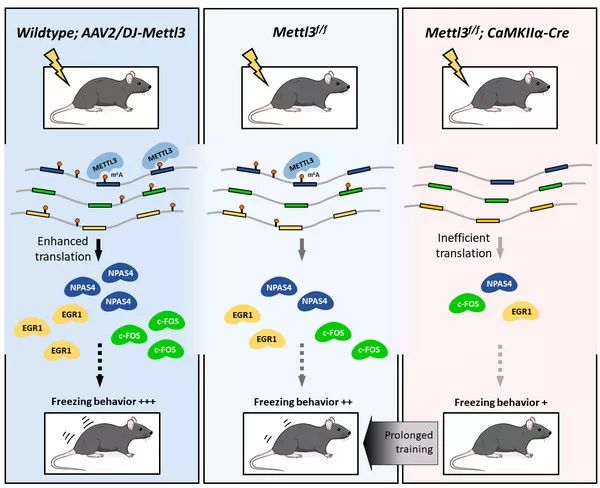
图:METTL3催化的 m6A 修饰增强长时记忆形成。增加学习训练强度可以弥补个体间 m6A 差异导致的学习能力差异。
来源:中国科学院遗传与发育生物学研究所
温馨提示:近期,微信公众号信息流改版。每个用户可以设置 常读订阅号,这些订阅号将以大卡片的形式展示。因此,如果不想错过“中科院之声”的文章,你一定要进行以下操作:进入“中科院之声”公众号 → 点击右上角的 ··· 菜单 → 选择「设为星标」







