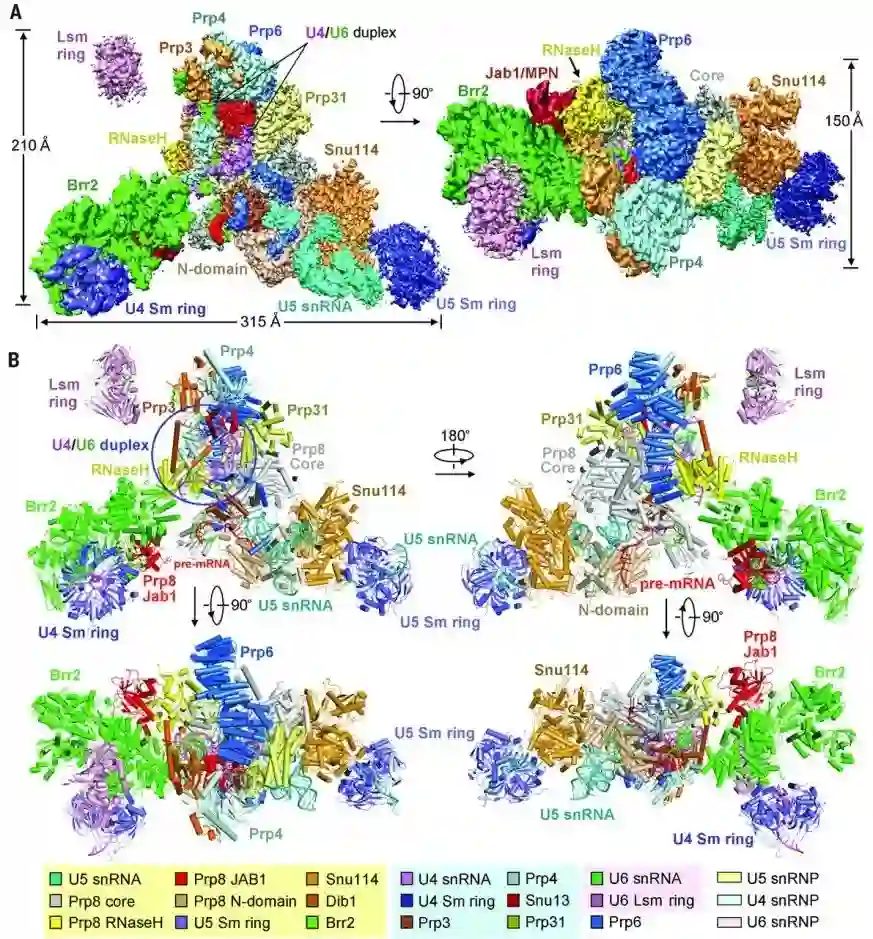
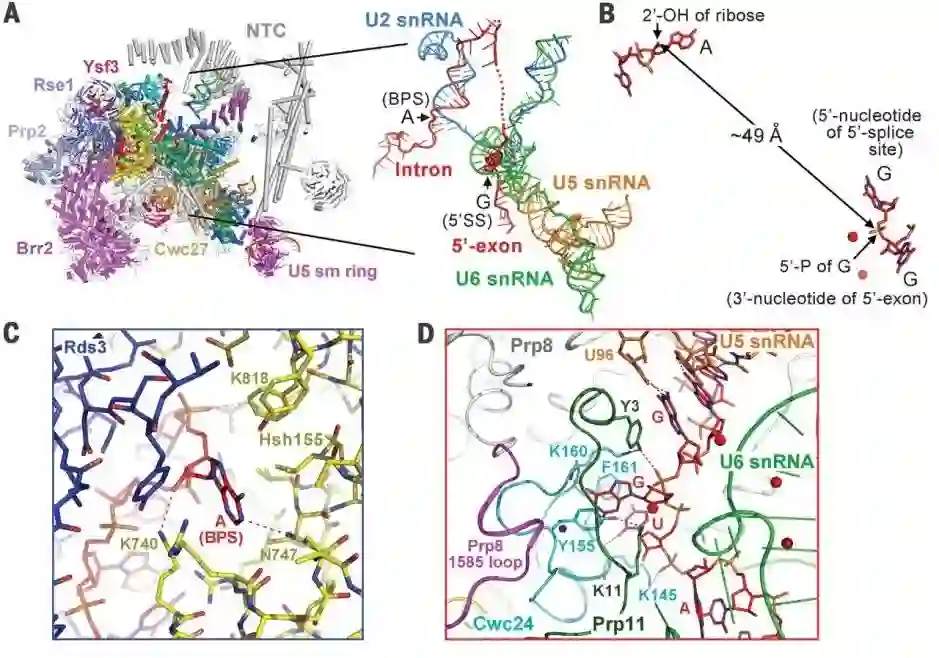
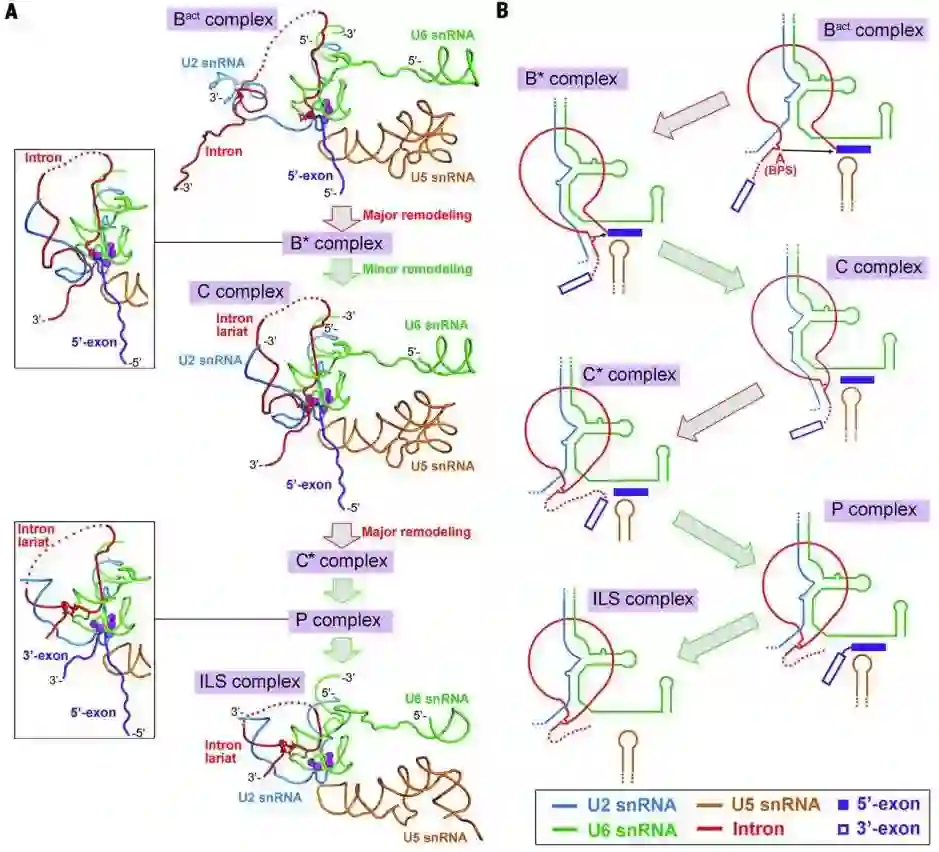
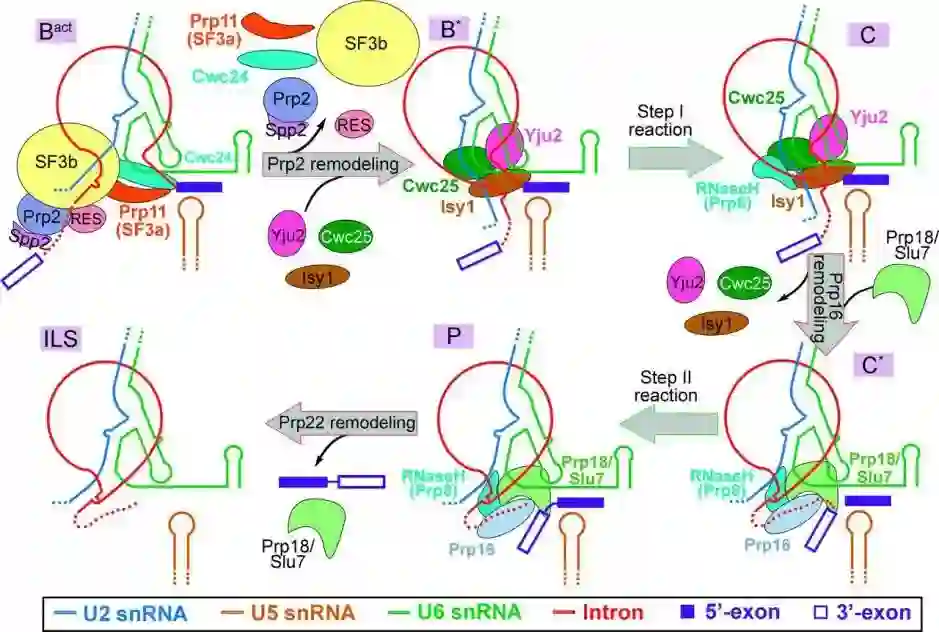
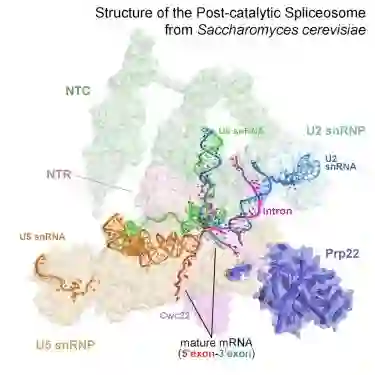
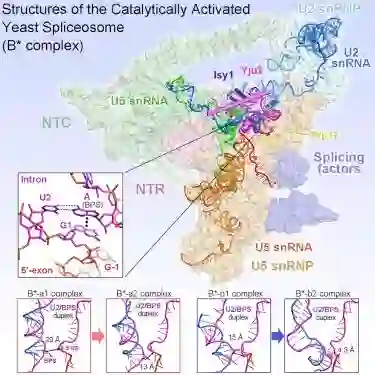
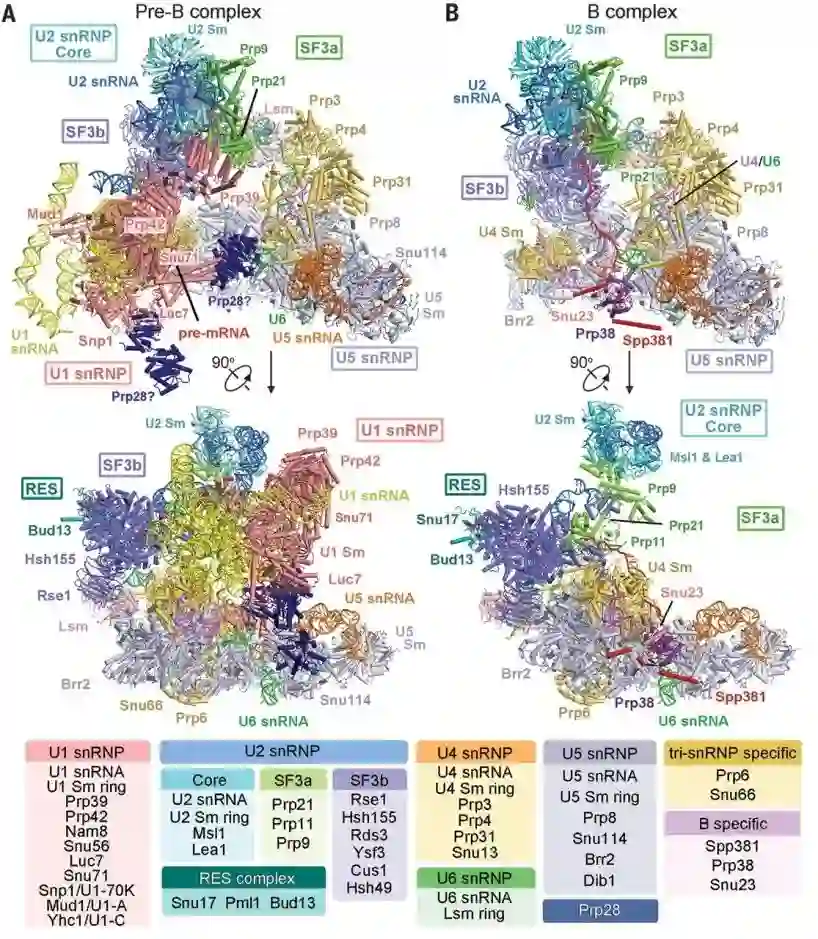
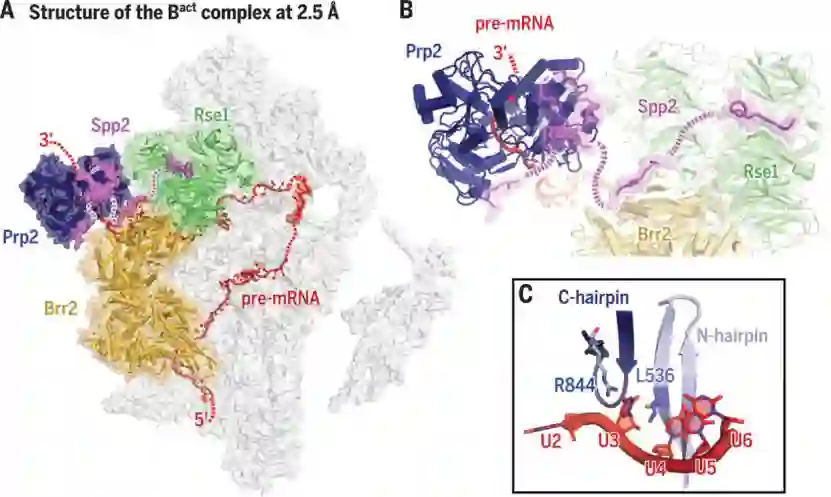
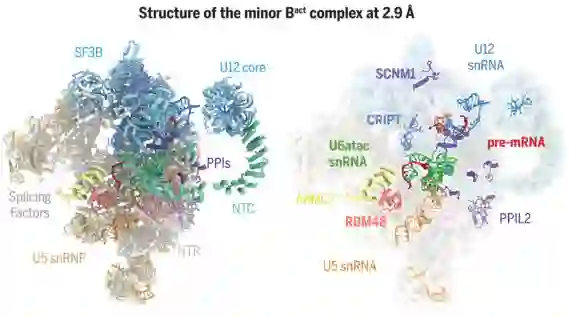
师从施一公,发表 6 篇Science/3篇 Cell,她被誉为「世界最具潜力女科学家」| 论文盘点









学术头条
新版微信更改了公众号推荐规则,不再以时间排序,而是以每位用户的阅读习惯为准进行算法推荐。在此情况下,学术头条和“学术菌”们的见面有如鹊桥相会一样难得(泪目)
那么,如果在不得不屈服于大数据的当下,你还想保留自己的阅读热忱,和学术头条建立长期的暧昧交流关系, 将学术头条纳入【星标】,茫茫人海中也定能相遇~

登录查看更多
相关内容

相关VIP内容
相关资讯